ГДЗ биология 6 класс Пасечник, Суматохин, Калинова Просвещение 2019-2020 Задание: 35 Выделение у растений и животных
Стр. 146. Вспомните
№ 1. Как растения удаляют ненужные вещества?
Накапливание продуктов обмена у растений происходит в вакуолях клеток, в специальных хранилищах, например, в млечных ходах у молочая или в смоляных ходах у хвойных. У многолетних растений для этого служат кора, а иногда древесина.
Удаление всех ненужных веществ у растений происходит через корни и опавшие листья. К осени в клетках листьев собирается большое количество вредных веществ, которые естественным путем удаляются из растения.
Через специальные железы в структуре растений выделяются в атмосферу летучие вещества, включая эфирные масла. Так происходит, например, у мяты, эвкалипта, мелиссы. Через устьица и чечевички коры у деревьев происходит выделение углекислого газа.
Выделение сахаров у растений происходит при помощи нектарников. У большинства представителей этого класса они находятся в цветках, но есть виды, у которых нектарники располагаются на листьях и стеблях.
№ 2. Как удаляются ненужные продукты жизнедеятельности у животных?
Все живые организмы в процессе своей жизнедеятельности образуют конечные продукты обмена, которые в дальнейшем выделяются в окружающую среду. Каждое животное стремится не только сохранить нужные ему питательные вещества, но также и удалить из своего организма результаты обмена, совершенно бесполезные, а иногда даже опасные для него. У разных животных этот процесс происходит по-разному.
Например, у амеб избавление от излишков воды происходит при помощи сократительной вакуоли, которая с определенной периодичностью сокращается и просто выталкивает собравшуюся в ней жидкость. Также через поверхность тела происходит удаление ненужных веществ у медуз и у гидр.
Функцию выведения у большинства многоклеточных животных выполняют специальные органы. Например, у дождевого червя для этого предназначены нефридии – специальные канальцы. У насекомых выделение происходит через трубчатые выросты кишечника. Основным органом выделения у всех позвоночных животных являются почки. У млекопитающих и птиц для выведения продуктов переработки в результате обмена веществ имеются потовые железы, кишечник. Также у рыб в процессе выведения углекислого газа участвуют жабры, у птиц и млекопитающих – лёгкие.
Стр. 149. Вопросы после параграфа
№ 1. Где у растений накапливаются продукты обмена веществ?
Продукты обмена веществ у растений накапливаются в клетках, а точнее – в их вакуолях, либо в специальных хранилищах – млечных и смоляных ходах. У многолетних растений накопление органических и неорганических веществ происходит в коре, либо в древесине. Также очень много вредных веществ, которые не нужны растению, собирается в листьях. За период цветения и плодоношения в них оседает много продуктов обмена, которые удаляются из растений в процессе листопада.
№ 2. Как происходит выделение вредных веществ у растений?
В отличие от животных, у растений отсутствует специальная выделительная система, поэтому вредные продукты обмена у них имеют свойство накапливаться в клетках и разных органах. Чаще всего они собираются в вакуолях клетках, специальных хранилищах. Например, в млечных ходах и в смоляных ходах, в коре многолетних деревьев и т.д.
За период созревания, цветения и плодоношения в растениях накапливается большое количество вредных веществ. Удаление их происходит как через корни, так и через листья, которые растения начинают сбрасывать к осени.
Выделение углекислого газа происходит через устьица и чечевички коры. Также различные летучие вещества, включая эфирные масла, выводятся и через специальные железы растений. А вот избавление от сахаров осуществляется через специальные образования – нектарники, которые находятся у большинства растений в цветках, но есть виды, у которых они есть на листьях и стеблях.
№3. Какие продукты обмена веществ выделяются из организма позвоночных животных через лёгкие, кишечник, потовые железы?
Из организма позвоночных животных через лёгкие выводится углекислый газ в процессе дыхания. Через кишечник происходит выведение непереваренных остатков пищи и вещества, которые не усваиваются организмом, в виде каловых масс. Выделение воды, соли и органических веществ происходит через потовые железы.
Стр. 149. Задание
№ 1. Выпишите из текста параграфа новые понятия, найдите их определения в интернет-источниках, энциклопедических словарях.
Новые понятия из текста параграфа:
Листопад – это биологический природный процесс, при котором в определенные периоды года растения массово сбрасывают листья в течение длительного промежутка времени.
Нектарники – это медовики или железы у растений, при помощи которых выделяется сахаристый сок или нектар, привлекающий насекомых или животных для опыления. Они очень разнообразны как по величине, так и по форме, происхождению, расположению на частях цветка. Чаще всего они возникают из эпидермальных и субэпидермальных клеток, которые многократно делятся, становятся меристематическими, образуя железы, разные по форме.
Нефридии – это выделительные органы у всех беспозвоночных животных, которые служат для осморегуляции, извлечения и дальнейшего выведения из их организма вредных продуктов в результате обмена веществ.
№ 2. Используя интернет-источники, научно-популярную литературу, подготовьте сообщение на тему «Значение процессов выделения у живых организмов».
В жизненноважных для любого живого организма процессах обмена всегда образуются конечные продукты, среди которых могут быть и бесполезные соединения, и вещества, которые могут быть вредными для его клеток. Например, в результате расщепления нуклеиновых кислот, аминокислот и прочих азотосодержащих соединений образуются такие вещества, как мочевина и мочевая кислота, аммиак, которые подлежат обязательному выведению из организмов. Также удалению подлежат и углекислый газ, который является результатом дыхания, и яды, которые попадают в организм вместе с воздухом, и излишки воды, гормонов и витаминов.
Как и все живые организмы на нашей планете, растения также выделяют активно или пассивно большое количество минеральных и органических веществ. У них процессы выделения происходят по-разному и через разные органы:
Вымывание дождевыми водами, которые стекают по стволам и листьям, значительное количество минеральных соединений;
Выделение кислорода, углекислого газа, солей, летучих метаболитов через устьица, чечевички коры, специальные железы и приспособления – нектарники;
Выделение через корни большого количества продуктов фотосинтеза, которые содержат целый ряд минеральных веществ, органические кислоты и сахара. Все они используются в дальнейшем симбиотическими организмами и микрофлорой ризосферы;
Сбрасывание листьев, в которых за время цветения и плодоношения происходит накопление балластных и ненужных растению веществ.
У представителей животного мира органы выделения в процессе эволюции стали появляться только на поздних стадиях. Например, у губок и кишечнополостных специальные выделительные органы отсутствуют, а выделение из организма конечных продуктов обмена происходит методом диффузии через поверхность всего тела. Впервые специальные органы для выделения, а именно протонефридии, появляются только у кольчатых червей. Далее у членистоногих, моллюсков и ланцетников органами выделения служат метанефридии, у насекомых и паукообразных – мальпигиевы сосуды, а у позвоночных хордовых животных – почки.
При всем разнообразии органов выделения у разных живых организмов в основе их функционирования лежит два основных процесса. Это ультрафильтрация и активный транспорт. При ультрафильтрации происходит прохождение жидкости под давлением через полупроницаемую мембрану, которая пропускает воду и низкомолекулярные растворенные в ней вещества, но в то же время удерживает белки и другие крупные молекулы. Благодаря активному транспорту осуществляется движение уже растворенных веществ против электрохимического и концентрационного градиентов, что связано с затратами энергии. В выделительных органах фильтрационная система дополнена еще и процессами активного транспорта. Значение полноценной работы выделительной системы в любом животном организме позволяет:
Контролировать объемы крови и прочих жидкостей внутренней среды;
Поддерживать кислотно-щелочное равновесие;
Обеспечивать постоянство осмотического давления крови и других жидкостей тела, а также ионного состава жидкостей внутренней среды организма;
Выводить все чужеродные вещества и конечные продукты азотистого обмена;
Стр. 149. Подумайте
Чем различается выделение веществ у растений и животных? Ответ представьте в виде таблицы.
§ 32. Выделение у растений и животных
1. Как растения удаляют ненужные вещества? 2. Как удаляются ненужные продукты жизнедеятельности у животных?
Организмы в процессе жизнедеятельности образуют конечные продукты обмена, которые выделяются в окружающую среду. Освобождение от них называют выделением. У растений и грибов, в отличие от животных, нет специальной выделительной системы. Продукты обмена у них могут накапливаться в клетках и органах. Например, плодовые тела старых шляпочных грибов содержат ядовитые вещества, поэтому их нельзя употреблять в пищу.
Выделение у растений
У растений продукты обмена веществ накапливаются в вакуолях клеток, в специальных хранилищах, например в смоляных ходах у хвойных, млечных ходах у одуванчика и молочая. У многолетних растений они накапливаются в коре, иногда в древесине. Удаление продуктов жизнедеятельности у растений происходит через корни и опавшие листья. Установлено, что к осени в клетках листьев накапливаются вредные для растения вещества, которые удаляются из растения вместе с опадающими листьями.
Через устьица и чечевички коры, например березы, из растения удаляется углекислый газ (см. рис. 69).
Выделение сахаров у растений осуществляется специальными образованиями — нектарниками. У большинства растений они находятся в цветках, а у некоторых — на стеблях и листьях. Нектар обладает бактерицидными свойствами и защищает завязь цветка от микроорганизмов. К тому же нектар наряду с окрашенным венчиком и ароматом цветков является важным приспособлением для привлечения насекомых, осуществляющих перекрёстное опыление.
Через специальные железы растений в атмосферу выделяются летучие вещества, в том числе эфирные масла. К эфиромасличным растениям относятся пеларгония, мята, мелисса, эвкалипт. Многие из них используются в лекарственных целях, а также для ароматизации продуктов, изготовления парфюмерной продукции.
Опавшие листья растений содержат неорганические и органические вещества и представляют собой очень ценное удобрение. Поэтому садоводы закладывают листья в компостные кучи. Благодаря опавшим листьям почва в лесу ежегодно обогащается перегноем. Вот почему их не надо жечь. Вполне понятно, что сбор опавших листьев и вообще удаление лесной подстилки в лесу отрицательно сказываются па жизни деревьев.
В городах, где почва и воздух загрязнены выхлопными газами автомобилей, выбросами промышленных предприятий, в листьях накапливаются ядовитые вещества. Поэтому их нельзя использовать для приготовления компоста, а почву следует регулярно удобрять.
У растений вредные продукты жизнедеятельности удаляются во время листопада. Листопад обычен у деревьев и кустарников. Изредка встречается у трав, например у крапивы, недотроги. Массовый листопад, приводящий к полной потере листьев, происходит у растений умеренного пояса с наступлением зимы, а у растений субтропиков и тропиков в засушливый период.
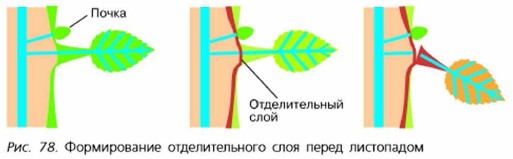
У древесных растений умеренных широт подготовка к листопаду начинается задолго до наступления морозов. Перед листопадом листья меняют свою окраску с зелёной на жёлтую, оранжевую, красную и др. (рис. 77).

Это связано с тем, что к осени происходит старение листьев. В них накапливаются продукты обмена веществ, разрушается зелёный пигмент — хлорофилл. Более стойкие пигменты (красные, жёлтые и др.) сохраняются и придают листьям осеннюю окраску.
Сигналом к наступлению листопада служит сокращение длины светового дня. Установлено, что деревья вблизи уличных фонарей сохраняют листья дольше, чем растущие вдали от них.
Опадение листьев связано с появлением у основания листа отделительного слоя из легко разъединяющихся клеток (рис. 78). Поэтому даже при небольшом ветре листья опадают. Продолжительность листопада у различных растений неодинаковая. Берёза сбрасывает листья около двух месяцев, липа и дуб — в течение двух недель. Деревья, растущие в одиночку или небольшими группами, где они подвержены ветру, теряют листья раньше, чем растущие в лесу. Листопадные деревья в лесах умеренных широт стоят без листьев до восьми-девяти месяцев в году, во влажных тропических лесах — иногда всего несколько дней. Листопад играет важную роль в жизни леса — опавшие листья перегнивают и служат хорошим удобрением, предохраняют корни от вымерзания.
Но не все растения сбрасывают листья. Некоторые сохраняют их всю зиму. Это вечнозелёные кустарнички: брусника, вереск, клюква. Мелкие, плотные листья этих растений, слабо испаряющие воду, сохраняются под снегом. С зелёными листьями зимует большинство хвойных деревьев и кустарников. Некоторые травы, например земляника, клевер, чистотел, тоже уходят под снег зелёными.
Выделение у животных
У животных в процессе обмена веществ также образуются продукты жизнедеятельности. Поэтому каждое животное стремится сохранять нужные вещества и удалять вещества, которые ему не нужны, а иногда и опасны для него.
Животные по-разному избавляются от продуктов обмена. Например, амёба от излишков воды избавляется с помощью сократительной вакуоли (рис. 79), которая, периодически сокращаясь, выталкивает наружу находящуюся в ней жидкость. Всей поверхностью тела удаляются ненужные вещества у гидр, медуз.
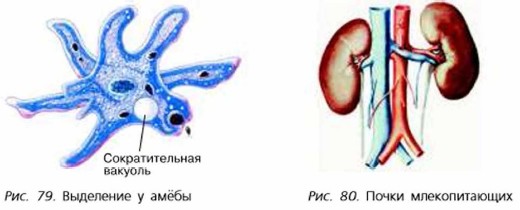
У большинства многоклеточных животных функцию выделения выполняют специальные органы. Так, у дождевого червя есть канальцы — нефридии (от греч. нефрос — почка). У насекомых функцию выделения выполняют трубчатые выросты кишечника. У рыб продукты обмена удаляются через жабры и почки, которые являются основным органом выделения у позвоночных животных (рис. 80). У птиц и млекопитающих в процессе удаления ненужных веществ участвуют также лёгкие, кишечник, потовые железы.
Выделение. Почки. Листопад
Ответьте на вопросы
1. Где у растений накапливаются продукты обмена веществ? 2. Как происходит выделение вредных веществ у растений? 3. Какие продукты обмена веществ выделяются из организма позвоночных животных через лёгкие, кишечник, потовые железы?
1. Выпишите из текста параграфа новые понятия, найдите их определения в интернет-источниках, энциклопедических словарях.
2. Используя интернет-источники, научно-популярную литературу, подготовьте сообщение на тему «Значение процессов выделения у живых организмов».
Чем различается выделение веществ у растений и животных? Составьте сравнительную таблицу.
Физиология растений. Обмен веществ, питание, рост растений
» data-shape=»round» data-use-links data-color-scheme=»normal» data-direction=»horizontal» data-services=»messenger,vkontakte,facebook,odnoklassniki,telegram,twitter,viber,whatsapp,moimir,lj,blogger»>
Физиология растений
Физиология растений изучает процессы, происходящие в организмах на разных уровнях организации: биоценотическом, организменном, органном, клеточном, субклеточном, молекулярном и даже субмолекулярном. В организме растения процессы всех уровней тесно взаимосвязаны. Изменение любого процесса отражается на всей жизнедеятельности организма. Сложность биологических исследований заключается еще и в том, что организм неотделим от среды, и все физиологические процессы тесно взаимосвязаны с условиями среды.
Любой физиологический процесс должен рассматриваться как результат эволюции, в течение которой выработалась способность растений к адаптации, приспособлению к изменяющимся условиям среды. Растительный организм непрерывно развивается в течение всей своей жизни. Это развитие разделено на определенные этапы, характеризующиеся специфическими признаками. Именно поэтому необходимо рассматривать растительный организм как непрерывно развивающуюся систему. В настоящее время применяют метод изучения таких систем от более простых к сложным уровням их организации. Этот подход позволяет проследить развитие отдельных физиологических процессов в целом растительном организме.
В физиологии растений используется вегетационный метод, основанный на исследовании выращиваемых экспериментальных растений в различных условиях опыта, широко применяют также методы биофизики и биохимии, методы культивирования клеток и тканей. Физиология – теоретическая основа клеточной и генетической (генной) инженерии.
Основные разделы физиологии растений посвящены таким процессам, как фотосинтез, транспорт веществ, дыхание, обмен веществ, почвенное питание, водный обмен, рост и развитие. Все эти процессы тесно связаны друг с другом и в живом организме неразделимы.
5.1. Обмен веществ
Организмы представляют собой открытые энергетические системы, непрерывно обменивающиеся с окружающей средой веществом и энергией. Метаболизм, или обмен веществ лежит в основе всех проявлений жизни. Различают внешний обмен – поглощение и выделение веществ, и внутренний обмен – химическое превращение этих веществ в клетке. Обмен веществ и поддержание целостности структуры любой живой системы требуют затраты определенной энергии и, следовательно, ее поступления извне. Первичным источником энергии у автотрофных организмов служит либо свет (у фототрофов), либо различные химические реакции (у хемотрофов).
Существование большинства живых организмов на Земле невозможно без использования запасенной энергии. Такая энергия накапливается в виде энергии химических связей углеводов, жиров и белков. Передатчиками энергии при ее поступлении и расходовании служат высокоэнергетические соединения типа АТФ, то есть аденозинтрифосфорной кислоты (аденозинтрифосфата).
В процессе обмена веществ строится тело растительного организма. Превращение чужеродных веществ в вещества собственного тела получило название ассимиляции. Ассимиляция всегда сопряжена с расходованием энергии.
Распад веществ, образующих организм, до более простых соединений называется диссимиляцией. При диссимиляции энергия высвобождается. Ассимиляция и диссимиляция представляют собой взаимосвязанные процессы обмена веществ и энергии в живых системах.
Помимо обмена веществ, происходящих в клетках, сами клетки обмениваются веществами с окружающей средой. Этот обмен происходит либо в виде свободного (пассивного) транспорта за счет энергии передвигающихся частиц в ходе диффузии и осмоса, либо в виде активного транспорта, при котором затрачивается определенная часть энергии, образующейся при диссимиляции. Другая ее часть расходуется на синтез структурных компонентов клетки и поддержание ее гомеостаза. Главнейшую роль в регуляции обмена веществ между клеткой и средой играет цитоплазматическая мембрана (плазмалемма), а в пределах клетки – эндоплазматическая сеть.
Основное количество используемой организмом энергии высвобождается в результате диссимиляции. В процесс диссимиляции вовлекаются запасные вещества клетки и всего организма. Известно 2 основных процесса диссимиляции: брожение и дыхание.
Брожение эволюционно более древний и энергетически менее выгодный процесс. В ходе брожения различные энергетически богатые субстраты (чаще всего углеводы) расщепляются до менее богатых соединений (спирта, масляной, молочной, уксусной кислот). Брожение характерно для многих прокариот и некоторых грибов. Например, процесс спиртового брожения суммарно можно выразить уравнением:
Из этого уравнения видно, что при сбраживании 1 молекулы сахара (глюкозы) образуется только 2 молекулы АТФ.
Дыхание энергетически более совершенно. В основе дыхания лежит биологическое окисление в так называемой цепи дыхания, содержащей специальные ферменты – оксиредуктазы. При полном окислении молекулы глюкозы до воды и диоксида углерода образуется 38 молекул АТФ:
Энергетически богатые субстраты в процессе дыхания окисляются до крайне бедных энергией соединений – воды и диоксида углерода. У большинства организмов в окислительно-восстановительных процессах активно используется кислород. Важнейшие этапы процесса дыхания у эукариотических организмов осуществляются в митохондриях. Интенсивность дыхания меняется в ходе развития растения. Сухие покоящиеся семена дышат слабо. При набухании и последующем прорастании семян интенсивность дыхания усиливается в сотни и тысячи раз. Самой высокой интенсивностью дыхания отличаются быстро растущие органы и ткани. С окончанием периода активного роста растений дыхание их тканей ослабевает, что связано с процессами старения протопласта.
Существует две формы ассимиляции: автотрофная и гетеротрофная. Автотрофная ассимиляция имеет огромное значение для живых существ, поскольку создает первичную продукцию, являющуюся основой всех цепей питания в экосистемах. При автотрофной ассимиляции неорганические вещества превращаются в органические. Этот процесс наиболее сложен. Гетеротрофная ассимиляция относительно проще, поскольку здесь происходит превращение одних органических веществ в другие. Она типична для большинства животных, грибов и части прокариот. Большинство растений и значительное число видов прокариот автотрофны.
Поскольку органические вещества представляют собой соединения углерода, решающее значение при создании первичной продукции имеет ассимиляция СО2. Это процесс восстановления, который ведет от максимально окисленного исходного вещества СО2 к менее окисленным продуктам, таким, как углеводы (СН2О)n. У растений и цианобактерий донором электронов, необходимых для восстановления углерода, служит вода, которая при отнятии электрона окисляется до кислорода. Такое преобразование энергии света называется аэробным (кислородным) фотосинтезом.
Реже, у фотобактерий, донором электронов выступают молекулярная сера или сероводород, водород или некоторые органические вещества. Кислород при таком процессе не выделяется. Такое преобразование энергии света в химическую энергию получило название анаэробного (бескислородного) фотосинтеза.
Относительно редко донорами электронов при автотрофной ассимиляции выступают различные неорганические соединения, например водород в метанообразующих бактериях, а энергия поступает в результате окисления сероводорода (Н2S), аммиака (NН3). Это процессы хемосинтеза. Солнечный свет для существования хемосинтезирующих организмов не нужен и все процессы могут протекать анаэробно.
5.2. Ассимиляция углерода (фотосинтез)
Органическое вещество состоит на 45% из углерода. Поэтому вопрос об источнике питания организмов углеродом чрезвычайно важен. Все организмы делят на автотрофные и гетеротрофные. Автотрофные организмы характеризуются способностью в качестве источника углерода использовать его минеральные формы, то есть синтезировать органическое вещество из неорганических соединений. Гетеротрофные организмы строят органическое вещество своего тела из уже имеющихся готовых органических соединений, то есть используют органические соединения как источник углерода. Для того, чтобы осуществить синтез органического вещества, необходима энергия. В зависимости от используемого соединения, а также от источников энергии, различают следующие основные типы питания углеродом и построения органических веществ.
Типы углеродного питания организмов
| Тип питания | Источник углерода | Источник водорода | Источник энергии |
| I. Гетеротрофный | Органическое вещество | Органическое вещество | Энергия окисления органических веществ |
| П. Автотрофный 1. Фотосинтез | СО2 | Н2О | Энергия света |
| 2.Бактериальный фотосинтез | СО2 | Н2S, Н2 и др. | Энергия света |
| 3. Хемосинтез | СО2 | Н2О, Н2S, Н2, NН3 | Энергия окисления неорганических веществ |
Из всех перечисленных типов питания углеродом фотосинтез зеленых растений, при котором построение органических соединений идет за счет простых неорганических веществ (СО2 и Н2О) с использованием энергии солнечного света, занимает совершенно особое место. Общее уравнение фотосинтеза:
Фотосинтез – это процесс, при котором энергия солнечного света превращается в химическую энергию. В самом общем виде это можно представить следующим образом: квант света (hv) поглощается хлорофиллом, молекула которого переходит в возбужденное состояние, при этом электрон переходит на более высокий энергетический уровень. В клетках фотоавтотрофов в процессе эволюции выработался механизм, при котором энергия электрона, возвращающегося на основной энергетический уровень, превращается в химическую энергию.
В процессе фотосинтеза из простых неорганических соединений (СО2, Н2О) строятся различные органические вещества. В результате происходит перестройка химических связей: вместо связей С–О и Н–О возникают связи С–С и С–Н, в которых электроны занимают более высокий энергетический уровень. Таким образом, богатые энергией органические вещества, которыми питаются и за счет которых получают энергию (в процессе дыхания) животные и человек, первоначально создаются в зеленом листе. Можно сказать, что практически вся живая материя на Земле является результатом фотосинтетической деятельности.
Почти весь кислород атмосферы фотосинтетического происхождения. Процессы дыхания и горения стали возможны только после того, как возник фотосинтез. Возникли аэробные организмы, способные усваивать кислород. На поверхности Земли процессы приняли биогеохимический характер, произошло окисление соединений железа, серы, марганца. Изменился состав атмосферы: содержание СО2 и аммиака снизилось, а кислорода и азота возросло. Возникновение озонового экрана, который задерживает опасную для живых организмов ультрафиолетовую радиацию, также является следствием появления кислорода.
Для того, чтобы процесс фотосинтеза протекал нормально, к хлоропластам должен поступать СО2. Основным поставщиком служит атмосфера, где количество СО2 составляет 0,03%. Для образования 1 г сахара необходимо 1,47 г СО2 – такое количество содержится в 2500 л воздуха.
Углекислый газ поступает в лист растения через устьица. Некоторое количество СО2 поступает непосредственно через кутикулу. При закрытых устьицах диффузия СО2 в лист резко сокращается.
Наиболее примитивная организация фотосинтетического аппарата у зеленых бактерий и цианобактерий. У этих организмов функцию фотосинтеза выполняют внутрицитоплазматические мембраны или особые структуры – хлоросомы, фикобилисомы. У водорослей уже эволюционно возникли органеллы (хроматофоры), в которых сосредоточены пигменты, они разнообразны по форме (спиральные, лентовидные, пластинчатые, звездчатые). Высшие растения характеризуются вполне сформировавшимся типом пластид в форме диска или двояковыпуклой линзы. Приняв форму диска, хлоропласты становятся универсальным аппаратом фотосинтеза. Фотосинтез протекает в зеленых пластидах – хлоропластах. В лейкопластах синтезируется и отлагается в запасной крахмал, в хромопластах накапливаются каротиноиды.
Размер дисковидных хлоропластов высших растений колеблется от 4 до 10 мкм. Число хлоропластов обычно составляет от 20 до 100 на клетку. Химический состав хлоропластов достаточно сложен и может быть охарактеризован следующими средними данными (% на сухую массу): белок – 35-55; липиды – 20-30; углеводы – 10; РНК – 2-3; ДНК – до 0,5; хлорофилл – 9; каротиноиды – 4,5.
В хлоропластах сосредоточены ферменты, принимающие участие в процессе фотосинтеза (окислительно-восстановительные, синтетазы, гидролазы). В хлоропластах, так же как и в митохондриях, имеется своя белоксинтезирующая система. Многие из ферментов, локализованных в хлоропластах, являются двухкомпонентными. Во многих случаях простетическая группа ферментов – это различные витамины. В хлоропластах сосредоточены многие витамины и их производные (витамины группы В, К, Е, D). В хлоропластах находится 80% Fe, 70% Zn, около 50% Сu от всего количества этих элементов в листе.
Хлоропласты окружены двойной мембраной. Толщина каждой мембраны 7,5-10 нм, расстояние между ними 10-30 нм. Внутреннее пространство хлоропластов заполнено бесцветным содержимым – стромой и пронизано мембранами. Мембраны, соединенные друг с другом, образуют плоские замкнутые полости (пузырьки) – тилакоиды (греч. «тилакоидес» – мешковидный). В хлоропластах содержатся тилакоиды двух типов. Короткие тилакоиды собраны в пачки и расположены друг над другом, напоминая стопку монет. Эти стопки называются гранами, а составляющие их тилакоиды – тилакоидами гран. Между гранами параллельно друг другу располагаются длинные тилакоиды – тилакоиды стромы. Между отдельными тилакоидами в стопках гран имеются узкие щели. Тилакоидные мембраны содержат большое количество белков, участвующих в фотосинтезе. В составе интегральных мембранных белков имеется много гидрофобных аминокислот. Это создает безводную среду и делает мембраны стабильнее.
Для того, чтобы световая энергия могла быть использована в процессе фотосинтеза, необходимо ее поглощение фоторецепторами – пигментами. Фотосинтетические пигменты – это вещества, которые поглощают свет определенной длины волны. Не поглощенные участки солнечного спектра отражаются, что и обусловливает окраску пигментов. Так, зеленый пигмент хлорофилл поглощает красные и синие лучи, тогда как зеленые лучи, в основном, отражаются. Видимая часть солнечного спектра включает длины волн от 400 до 700 нм.
Состав пигментов зависит от систематического положения группы организмов. У фотосинтезирующих бактерий и водорослей пигментный состав разнообразен (хлорофиллы, бактериохлорофиллы, бактериородопсин, каротиноиды, фикобилины). Их набор и соотношение специфичны для различных групп организмов. Пигменты, сконцентрированные в пластидах, можно разделить на три группы: хлорофиллы, каротиноиды, фикобилины.
Важнейшую роль в процессе фотосинтеза играют зеленые пигменты –хлорофиллы. Французские ученые П.Ж. Пелетье и Ж. Кавенту (1818) выделили из листьев зеленое вещество и назвали его хлорофиллом (от греч. «хлорос» – зеленый и «филлон» – лист). В настоящее время известно около десяти хлорофиллов. Они отличаются по химическому строению, окраске, распространению среди групп организмов. У всех высших растений содержатся хлорофиллы a и b. Хлорофилл c обнаружен в диатомовых водорослях, хлорофилл d – в красных водорослях. Кроме того, известны бактериохлорофиллы (а, b, c, d), содержащиеся в клетках фотосинтезирующих бактерий. В клетках зеленых бактерий имеются бактериохлорофиллы с и d, в клетках пурпурных бактерий – бактериохлорофиллы a и b. Основными пигментами, без которых фотосинтез не идет, являются хлорофилл a для зеленых высших растений и водорослей, и бактериохлорофиллы – для бактерий.
Впервые точное представление о пигментах зеленого листа высших растений было получено благодаря работам крупнейшего российского ботаника М.С. Цвета (1872-1919). Он разработал новый хроматографический метод разделения веществ и выделил пигменты листа в чистом виде. Оказалось, что листья высших растений содержат хлорофилл a и хлорофилл b, а также каротиноиды (каротин, ксантофилл). Хлорофиллы, так же, как и каротиноиды, нерастворимы в воде, но хорошо растворимы в органических растворителях. Хлорофиллы a и b различаются по цвету: хлорофилл a имеет сине-зеленый оттенок, хлорофилл b – желто-зеленый. Содержание хлорофилла a в листе примерно в 3 раза больше по сравнению с хлорофиллом b. По химическому строению хлорофиллы – сложные эфиры дикарбоновой органической кислоты – хлорофиллина и двух остатков спиртов – фитола (С20Н39ОН) и метилового (СН3ОН). Эмпирическая формула хлорофилла С55Н72О5N4Мg (рис. 5.1).
Органическая дикарбоновая кислота хлорофиллин представляет собой азотсодержащее металлорганическое соединение, относящееся к магнийпорфиринам: (СООН) 2 = С32Н30ОN4Мg.
В хлорофилле водород карбоксильных групп замещен остатками двух спиртов – метилового СН3ОН и фитола С20Н39ОН, поэтому хлорофилл является сложным эфиром.

Рис. 5.1. Структурная формула хлорофилла а.
Хлорофилл b отличается тем, что содержит на два атома водорода меньше и на один атом кислорода больше (вместо группы СН3 группа СНО). В связи с этим, молекулярная масса хлорофилла a – 893 и хлорофилла b – 907.
В центре молекулы хлорофилла расположен атом магния, который соединен с четырьмя атомами азота пиррольных группировок. В пиррольных группировках хлорофилла имеется система чередующихся двойных и простых связей. Это хромофорная группа хлорофилла, обусловливающая поглощение определенных лучей солнечного спектра и его окраску.
Еще К.А. Тимирязев обратил внимание на близость химического строения двух важнейших пигментов: зеленого – хлорофилла листьев и красного – гемина крови. Действительно, если хлорофилл относится к магнийпорфиринам, то гемин – к железопорфиринам. Сходство это служит еще одним доказательством единства всего органического мира.
Молекула хлорофилла полярна, ее порфириновое ядро обладает гидрофильными свойствами, а фитольный конец – гидрофобными. Это свойство молекулы хлорофилла обусловливает определенное расположение ее в мембранах хлоропластов. Порфириновая часть молекулы связана с белком, а фитольная цепь погружена в липидный слой.
Хлорофилл способен к избирательному поглощению света. Спектр поглощения определяется его способностью поглощать свет определенной длины волны (определенного цвета). Для того чтобы получить спектр поглощения, К.А. Тимирязев пропускал луч света через раствор хлорофилла. Было показано, что хлорофилл в той же концентрации, как в листе, имеет две основные линии поглощения в красных и сине-фиолетовых лучах. При этом хлорофилл a в растворе имеет максимум поглощения 429 и 660 нм, тогда как хлорофилл b – при 453 и 642 нм (рис. 5.2).

Рис. 5.2. Спектры поглощения хлорофилла а и хлорофилла b
Наряду с зелеными пигментами в хлоропластах и хроматофорах содержатся пигменты, относящиеся к группе каротиноидов. Каротиноиды – это желтые и оранжевые пигменты алифатического строения, производные изопрена. Каротиноиды содержатся во всех высших растениях и у многих микроорганизмов. Это самые распространенные пигменты с разнообразными функциями. Каротиноиды, содержащие кислород, получили название ксантофиллы. Основными представителями каротиноидов у высших растений являются два пигмента – бета-каротин (оранжевый) С40Н56 и ксантофилл (желтый) С40Н56О2. Каротин состоит из 8 изопреновых остатков. При разрыве углеродной цепочки пополам и образовании на конце спиртовой группы каротин превращается в 2 молекулы витамина А.
Бета-каротин имеет два максимума поглощения, соответствующие длинам волн 482 и 452 нм. В отличие от хлорофиллов каротиноиды не поглощают красные лучи, а также не обладают способностью к флуоресценции. Подобно хлорофиллу каротиноиды в хлоропластах и хроматофорах находятся в виде нерастворимых в воде комплексов с белками. Каротиноиды всегда присутствуют в хлоропластах, они принимают участие в процессе фотосинтеза. Поглощая световую энергию в определенных участках солнечного спектра, они передают энергию этих лучей на молекулы хлорофилла. Тем самым, они способствуют использованию лучей, которые хлорофиллом не поглощаются. Физиологическая роль каротиноидов не ограничивается их участием в передаче энергии на молекулы хлорофилла. Каротиноиды выполняют защитную функцию, предохраняя молекулы хлорофилла от разрушения на свету в процессе фотоокисления (рис. 5.3).

Рис. 5.3. Структурная формула бета-каротина
Фикобилины – красные и синие пигменты, содержащиеся у цианобактерий и красных водорослей. В основе химического строения фикобилинов лежат 4 пиррольные группировки. В отличие от хлорофилла у фикобилинов пиррольные группы расположены в виде открытой цепочки (рис. 5.4).

Рис. 5.4. Структурная формула хромофорной группы фикоэритринов
Фикобилины представлены пигментами: фикоцианином, фикоэритрином и аллофикоцианином. Фикоэритрин – это окисленный фикоцианин. Красные водоросли, в основном, содержат фикоэритрин, а цианобактерии – фикоцианин. Фикобилины образуют прочные соединения с белками (фикобилинпротеиды). В отличие от хлорофиллов и каротиноидов, расположенных в мембранах, фикобилины концентрируются в особых гранулах (фикобилисомах), тесно связанных с мембранами тилакоидов. Фикобилины поглощают лучи в зеленой и желтой частях солнечного спектра. Это та часть спектра, которая находится между двумя основными линиями поглощения хлорофилла. Фикоэритрин поглощает лучи с длиной волны 495-565 нм, а фикоцианин – 550-615 нм. Сравнение спектров поглощения фикобилинов со спектральным составом света, в котором проходит фотосинтез у цианобактерий и красных водорослей, показывает, что они очень близки. Это позволяет считать, что фикобилины поглощают энергию света и, подобно каротиноидам, передают ее на молекулу хлорофилла, после чего она используется в процессе фотосинтеза. Наличие фикобилинов у водорослей является примером приспособления организмов в процессе эволюции к использованию участков солнечного спектра, которые проникают сквозь толщу морской воды (хроматическая адаптация).
Фотосинтез – это сложный многоступенчатый окислительно-восстановительный процесс, в котором происходит восстановление углекислого газа до углеводов и окисление воды до кислорода. В процессе фотосинтеза происходят не только реакции, идущие с использованием энергии света, но и темновые, не требующие непосредственного участия энергии света. Можно привести следующее доказательство существования темновых реакций в процессе фотосинтеза: фотосинтез ускоряется с повышением температуры. Отсюда прямо следует, что какие-то этапы этого процесса непосредственно не связаны с использованием энергии света. Процесс фотосинтеза включает следующие этапы: 1) фотофизический; 2) фотохимический (световой); 3) ферментативный (темновой).
Согласно законам фотохимии, при поглощении кванта света атомом или молекулой какого-либо вещества электрон переходит на другую, более удаленную орбиталь, то есть на более высокий энергетический уровень (рис. 5.5).

Рис. 5.5. Переходы между возбужденными состояниями хлорофилла после поглощения квантов синего и красного света
Наибольшей энергией обладает электрон, отдаленный от ядра атома и находящийся на достаточно большом расстоянии от него. Каждый электрон переходит на более высокий энергетический уровень под влиянием одного кванта света, если энергия этого кванта равна разнице между этими энергетическими уровнями. Все фотосинтезирующие организмы содержат какой-либо тип хлорофилла. В молекуле хлорофилла два уровня возбуждения. Именно с этим связано и то, что он имеет две основные линии поглощения. Первый уровень возбуждения обусловлен переходом на более высокий энергетический уровень электрона в системе сопряженных двойных связей, а второй – с возбуждением неспаренных электронов атомов азота и кислорода в порфириновом ядре. При поглощении света электроны переходят в колебательное движение и перемещаются на следующие орбитали с более высоким энергетическим уровнем.
Наиболее высокий энергетический уровень – это второй синглетный уровень. Электрон переходит на него под влиянием сине-фиолетовых лучей, кванты которых содержат больше энергии.
Из возбужденного, первого синглетного и триплетного состояния молекула хлорофилла также может переходить в основное. При этом ее дезактивация (потеря энергии) может проходить:
1) путем выделения энергии в виде света (флуоресценция и фосфоресценция) или в виде тепла;
2) путем переноса энергии на другую молекулу пигмента;
3) путем затрачивания энергии на фотохимические процессы (потеря электрона и присоединение его к акцептору).
В любом из указанных случаев молекула пигмента дезактивируется и переходит на основной энергетический уровень.
Хлорофилл имеет две функции – поглощение и передачу энергии. При этом основная часть молекул хлорофилла – более 90% всего хлорофилла хлоропластов входит в состав светособирающего комплекса (ССК). Светособирающий комплекс выполняет роль антенны, которая эффективно поглощает свет и переносит энергию возбуждения к реакционному центру. Кроме большого числа (до нескольких сотен) молекул хлорофилла ССК содержит каротиноиды, а у некоторых водорослей и цианобактерий – фикобилины, которые увеличивают эффективность усвоения света.
В процессе эволюции в растениях выработался механизм, позволяющий наиболее полно использовать кванты света, падающие на лист подобно каплям дождя. Механизм этот заключается в том, что энергия квантов света улавливается 200-400 молекулами хлорофилла и каротиноидами ССК и передается одной молекуле – реакционному центру. Расчеты показали, что в одном хлоропласте до 1 млрд молекул хлорофилла. Теневыносливые растения имеют, как правило, больший размер ССК по сравнению с растениями, растущими в условиях высокой освещенности. В реакционных центрах в результате фотохимических реакций образуются первичные восстановитель и окислитель. Они затем вызывают цепь последовательных окислительно-восстановительных реакций. В итоге энергия запасается в виде восстановленного никотинамидадениндинуклеотидфосфата (НАДФ•Н+) и аденозинтрифосфата (АТФ), который синтезируется из аденозиндифосфата (АДФ) и неорганической фосфорной кислоты за счет реакции фотосинтетического фосфорилирования. Следовательно, НАДФ•Н+ и АТФ – основные продукты световой фазы фотосинтеза. Таким образом, в первичных процессах фотосинтеза, связанных с поглощением молекулой хлорофилла кванта света, важную роль играют процессы передачи энергии. Фотофизический этап фотосинтеза заключается в том, что кванты света поглощаются и переводят молекулы пигментов в возбужденное состояние. Затем эта энергия переносится на реакционный центр, осуществляющий первичные фотохимические реакции: разделение зарядов. Дальнейшее превращение энергии света в химическую энергию проходит ряд этапов, начиная с окислительно-восстановительных превращений хлорофилла и включая как фотохимические (световые), так и энзиматические (темновые) реакции.
То есть фотосинтез включает преобразование энергии (явление, получившее название светового процесса) и превращение вещества (темновой процесс). Световой процесс происходит в тилакоидах, темновой – в строме хлоропластов. Два процесса фотосинтеза выражаются отдельными уравнениями:
12Н2О =12Н2 + 6О2 + энергия АТФ (световой процесс).
Из этого уравнения видно, что кислород, выделяемый при фотосинтезе, образуется при разложении молекул воды. Кроме того, световая энергия используется на синтез аденозинтрифосфорной кислоты (АТФ) в ходе фотофосфорилирования.
В темновых реакциях используются продукты, накопленные в световой фазе. Суть темновых реакций сводится к фиксации СО2 и включению его в молекулу сахара. Этот процесс получил название цикла Кальвина по имени американского биохимика, подробно изучившего последовательность темновых реакций. Использование воды в качестве источника водорода для синтеза органических молекул дало растениям в процессе эволюции большое преимущество в силу повсеместного ее присутствия (вода является самым распространенным минералом на Земле).
Поскольку весь кислород фотосинтеза выделяется из воды, итоговое уравнение принимает вид:
Вода в правой части уравнения не подлежит сокращению, так как ее кислород имеет иное происхождение (из СО2). Следовательно, фотосинтез – это окислительно-восстановительный процесс, в котором вода окисляется до молекуляргого кислорода (О2), а углекислый газ восстанавливается водородом воды до углеводов.
По завершении каждого цикла образуется конечный продукт: одна молекула сахара, который ложится в основу первичного органического вещества, образующегося при фотосинтезе.
5.3. Минеральное питание
В тело растения многие химические элементы поступают в форме ионов, диссоциированных в водных растворах, через корень, с помощью которого осуществляется минеральное, или почвенное, питание. Минеральное питание – это совокупность процессов поглощения из почвы, передвижения и усвоения химических биогенных элементов, то есть элементов, необходимых для жизни растительных организмов. Питательные вещества представляют собой доступные для растений минеральные соединения, в которых содержатся эти элементы. В особенно больших количествах растениям необходимы макроэлементы N, S, P, K, Mg, Ca. Напротив, в крайне малых количествах необходимы такие микроэлементы, как B, Mn, Cu, Zn, Mo, Co.
Азот и сера – важные структурные элементы органических соединений, например белков и нуклеиновых кислот. При недостатке азота сокращается синтез белков (в том числе ферментов), что проявляется в отставании роста, хлорозе (пожелтении) листьев. В большинстве природных экосистем именно азот находится в так называемом главном минимуме, то есть его недостаток ограничивает рост растений. Соединения фосфора играют главнейшую роль в энергетическом обмене. Калий, магний и кальций влияют на гидратацию коллоидов протопласта. При недостатке кальция особенно сильно повреждаются меристемы, при недостатке магния также возникает хлороз. Микроэлементы действуют как составные части ферментов.
Некоторые ионы являются антагонистами. Известен антагонизм одно- и двухвалентных катионов (соответственно K, Na по отношению к Mg, Ca). Если при повышенных концентрациях ионы K и Na ядовиты для растения, то их токсичное действие возможно частично нейтрализовать добавлением в почву определенных доз ионов-антагонистов (Mg и Ca).
Механизм поступления ионов в корень сложен. Он связан с их адсорбцией и активным поглощением из почвы, при этом затрачивается энергия. Проникнув в корень, питательные вещества далее перемещаются по тканям растения. Существуют два основных направления перемещений больших количеств веществ в растении: транспирационный (или восходящий) ток, то есть транспорт воды и растворенных в ней веществ от корня к побегам, и ток ассимилятов (нисходящий), то есть передвижение выработанных при фотосинтезе веществ из листьев к частям растения (стеблям, корню, репродуктивным органам – цветкам, плодам, семенам). Для таких перемещений, получивших название дальнего транспорта, есть особые проводящие ткани – ксилема и флоэма. Ближний транспорт к проводящим тканям и от них осуществляется через клетки других тканей тремя путями: по симпласту, то есть по соединенным между собой через плазмодесмы протопластам клеток, по апопласту, то есть по взаимосвязанной системе клеточных стенок и межклетников, и по системе вакуолей. Симпласт служит для транспорта минеральных и органических веществ, апопласт – только для воды и растворенных в ней ионов минеральных соединений, система вакуолей – для воды.
Главный путь дальнего транспорта воды и ионов – транспирационный ток по ксилеме. Для ближнего транспорта в корне используются симпласт и апопласт. Главным фильтрующим барьером на пути ионов в корне оказывается эндодерма с ее поясками Каспари. Через эндодерму ионы проникают по пропускным клеткам путем активного транспорта. Переход ионов в сосуды или трахеиды ксилемы может быть пассивным или активным. В стебле ионы активно извлекаются из сосудов и транспортируются в горизонтальном направлении, в основном по паренхиме сердцевинных лучей. В листе из окончаний сосудов ионы в водных растворах выходят пассивно вместе с транспирационным током и в течение вегетационного периода могут накапливаться в большом количестве по мере испарения воды. Часть накопленных ионов удаляется вместе с осенним листопадом (Са и Мg), другая часть отводится из листьев.
Основным двигателем транспирационного тока является транспирация, то есть выделение водяного пара в атмосферу, а также гуттация – выделение капельно жидкой воды водяными устьицами – гидатодами.
Основное количество воды испаряется листьями через устьица (устъичная транспирация). Пары воды по межклетникам мезофилла листа попадают в устьичные полости и через устьичные щели испаряются в атмосферу. Движения устьиц (открывание и закрывание) регулируют интенсивность транспирации. Малую часть воды растение теряет через кутикулу (кутикулярная транспирация), но ее интенсивность в 10-20 раз ниже устьичной. В теле растения происходит непрерывный ток воды из почвы в атмосферу. Присасывающее действие поднимает столб воды, заполняющий просветы сосудов со скоростью от 1 до 100 м/ч. Кроме того, при подъеме воды по сосудам важную роль играет сила сцепления (когезия) между молекулами воды и адгезия – сила сцепления молекул воды со стенками сосудов ксилемы. Создаются условия, позволяющие удерживать в сосудах столб воды высотой до 140 м. Высота самых больших деревьев, видно, не может превысить эту величину.
В процессе жизнедеятельности растения накапливают первичные и вторичные метаболиты. Накопление метаболитов определяется тремя главными причинами: 1. необходимостью создания резерва энергетических и пластических веществ, 2. защиты от поедания животными и внедрения болезнетворных организмов и, наконец, 3. необходимостью концентрации отбросов (шлаков), подлежащих удалению из организма.
К резервным энергетическим и пластическим веществам обычно относятся лишь первичные метаболиты (белки, жиры, углеводы). Перед листопадом или завяданием надземных частей многолетних растений резервные вещества оттягиваются из листьев в подземные органы. У однолетников они концентрируются в семенах. Вторичные метаболиты (терпеноиды, алкалоиды, полифенольные соединения) синтезируются в клетках различных органов. Чаще всего они выполняют защитную роль, хотя могут вовлекаться вновь в основной обмен веществ (реутилизироваться). Изредка вторичные метаболиты оказываются в роли шлаков и удаляются при опаде тех частей растения, где они накапливаются, либо выделяются при разрыве соответствующих структур выделительных тканей. Распространенным отбросом в растении выступает оксалат кальция (щавелевокислый кальций), в который превращаются отходы как первичного, так и вторичного метаболизма. Обычно он удаляется при листопаде вместе с опадающими листьями или отслаивающейся корой (коркой) стеблей.
Транспорт ассимилятов (дальний и ближний) происходит с затратой энергии по симпласту, то есть по системе протопластов живых клеток. Количество продуктов, образовавшихся за день в процессе фотосинтеза, превосходит массу хлоропластов во много раз. В этой связи большое значение имеет отток ассимилятов в другие части клетки, то есть внутриклеточный транспорт. Проникновение через мембрану хлоропластов крахмала, сахарозы и фосфорилированных гексоз затруднено. Наиболее легко через мембраны хлоропластов проникают триозофосфаты: фосфоглицериновый альдегид (ФГА), фосфодиоксиацетон (ФДА). Предполагается, что образующиеся в хлоропластах более сложные углеводы распадаются на триозофосфаты и в таком виде передвигаются в цитоплазму, где могут служить материалом для ресинтеза гексоз, сахарозы, крахмала. Благодаря указанным превращениям концентрация триозофосфатов в цитоплазме не снижается, что способствует их притоку по градиенту концентрации. Отток триозофосфатов сопровождается переносом ортофосфата внутрь хлоропласта с помощью переносчика (транспортные белки). Это позволяет сохранять в хлоропластах содержание фосфора, необходимого для синтеза АТФ, на постоянном уровне. Наряду с превращением в сахарозу, поступившие в цитоплазму триозофосфаты участвуют в челночных превращениях. ФДА окисляется до фосфоглицериновой кислоты (3-ФГК) через дифосфоглицериновую кислоту (1,3-ДФГК). Это сопряжено с накоплением энергии в форме НАДН и АТФ. Именно этим путем АТФ и НАДН переносятся из хлоропласта в цитоплазму. При этом молекула 3-ФГК возвращается обратно в хлоропласт. Образовавшиеся в хлоропластах белки также распадаются и оттекают в цитоплазму в виде аминокислот. На свету проницаемость мембран хлоропластов повышается, что способствует оттоку из них различных веществ. Поступившие в цитоплазму органические соединения используются клеткой и направленно передвигаются к ситовидным трубкам. В зависимости от густоты проводящих элементов в листе расстояния от хлоренхимной клетки, производящей ассимиляты, до ситовидных элементов флоэмы могут быть различны. Однако в среднем оно не превышает 3-4 клеток и составляет сотые доли миллиметра.
Межклеточный паренхимный транспорт может осуществляться как по симпласту, так и частично по клеточным стенкам. Скорость перемещения ассимилятов в паренхимных тканях равняется примерно 10-60 см/ч, это заметно выше скорости обычной диффузии. При передвижении веществ по плазмодесмам такая скорость может достигаться лишь при большой дополнительной затрате энергии. Вместе с тем не у всех растений хорошо развиты плазмодесмы между клетками мезофилла и флоэмы. Показано, что клетки мезофилла листа обладают значительной секреторной способностью и легко выделяют сахарозу в свободное пространство. Сахароза на пути к ситовидным трубкам передвигается по апопласту. Соотношение апопластного или симпластного пути зависит от вида растения, транспортируемого сахара, типа сопутствующих клеток, числа плазмодесм. По апопласту главным образом передвигается сахароза, при этом отмечается малое количество плазмодесм в клетках флоэмы. По симпласту транспортируется не только сахароза, но и другие сахара (рафиноза), а в клетках флоэмы наблюдается много плазмодесм. У одних растений преобладает апопластный транспорт, у других – симпластный.
Ассимиляты (глюкоза) из мест их образования проходят по симпласту или апопласту до клеток-спутниц и окружающих ситовидные трубки паренхимных клеток. В этих клетках сначала происходит превращение глюкозы в сахарозу, которая затем в результате активного переноса (с затратой энергии и расходования АТФ) транспортируется в ситовидные трубки. Дальний транспорт сахарозы осуществляется по ситовидным трубкам. В местах потребления ассимилятов или отложения запасных веществ сахароза переходит из ситовидных трубок и достигает места назначения по симпластическому пути опять уже в виде глюкозы.
Растущие органы и ткани (листья, меристемы, корневые окончания, плоды) активно притягивают к себе ассимиляты из мест их хранения, а также из закончивших рост листьев.
5.4. Рост и развитие растений
В основе роста и развития целого организма и отдельной клетки лежит обмен веществ. В процессе жизни каждого организма происходят постоянные качественные и количественные изменения, прерываемые периодами покоя. Необратимое количественное увеличение структур, объема и массы живого тела и его частей получило название роста. Развитие – это качественные изменения организма. Рост и развитие тесно связаны между собой, оба процесса регулируются на клеточном уровне. Рост органов и всего организма слагается из роста его клеток. Основные этапы роста, а также и развития на клеточном уровне – деление клеток и их растяжение, то есть увеличение клеточного потомства и увеличение их размеров. В многоклеточных организмах одним из показателей роста будет увеличение числа клеток в результате клеточного деления. Растительная клетка способна к росту растяжением, чему содействуют особенности строения ее оболочки. Особенности роста различны у разных систематических групп организмов. У высших растений рост тесно связан с деятельностью меристем. Рост, так же как и развитие, контролируется фитогормонами – химическими соединениями, вырабатываемыми в малых количествах, но способных давать значительный физиологический эффект. Фитогормоны, выработанные в одной части растения, транспортируются в другую часть, вызывая там соответствующие изменения в зависимости от генной модели воспринимающей клетки.
Известны три класса фитогормонов, действующих, по преимуществу, как стимуляторы: ауксины (индолилуксусная, нафтилуксусная кислоты) (рис. 5.6), цитокинины (кинетин, зеатин) (рис. 5.7) и гиббереллины (С10 – гибериллин).
Два класса гормонов (абсцизовая кислота и этилен) оказывают тормозящее действие (рис. 5.8).
Заметное воздействие на рост и развитие растений оказывают ведущие факторы среды: свет, тепло и влага. Комплекс факторов и фитогормонов действует либо независимо, либо взаимодействуя друг с другом.


Рис. 5.7. Структурные формулы цитокининов

Рис. 5.8. Структурная формула абсцизовой кислоты
Интенсивность роста существенным образом связана с питанием растений, особенно с азотным и фосфорным. Типы роста различных органов определяются характером расположения меристем. Стебли и корни растут верхушками, они имеют апикальный рост. Зона нарастания листьев часто находится у их основания и они имеют базальный рост. Характер роста органа зависит от видовой специфичности. У злаков, например, рост стебля осуществляется у основания междоузлий, преобладает интеркалярный рост. Важная особенность роста растений – его ритмичность (чередование процессов интенсивного и замедленного роста). Она зависит не только от изменений внешних факторов среды, но и контролируется внутренними факторами (эндогенно), закрепленными в процессе эволюции. В целом рост растения складывается из четырех фаз: начальной, интенсивного роста, замедления роста и стационарного состояния. Это связано с особенностями различных стадий онтогенеза (индивидуального развития) растений. Так, переход растения к репродуктивному состоянию обычно сопровождается ослаблением активности меристем. Процессы роста могут прерываться продолжительными периодами торможения, наступление которых в северных широтах связано с концом лета и приближением зимы. Иногда у растений наблюдается как бы остановка роста – состояние покоя. Покой у растений – это такое физиологическое состояние, при котором резко снижаются скорость роста и интенсивность обмена веществ. Оно возникло в ходе эволюции как приспособление для переживания неблагоприятных условий среды в разные периоды жизненного цикла или сезона года. Покоящееся растение устойчиво к морозам, жаре, засухе. В состоянии покоя могут находиться растения (зимой, во время засухи), их семена, почки, клубни, корневища, луковицы, споры. Семена многих растений способны к длительному покою, обусловливающему их длительную сохранность в почве. Известен случай вызревания растения из семени одного из бобовых, пролежавшего в условиях вечной мерзлоты 10000 лет. В состоянии покоя находятся, например, клубни картофеля, благодаря чему они долго не прорастают. В понятие «развитие» вкладываются два смысла: индивидуальное развитие отдельного организма (онтогенез) и развитие организмов в ходе эволюции (филогенез). Физиология растений занимается изучением, главным образом, развития в онтогенезе.
Меристематические клетки тотипотентны (омнипотентны) – любая живая клетка может дать начало недифференцированным клеткам, способным развиваться самыми различными путями (рис. 5.9). Переход меристематической клетки к росту сопровождается появлением в ней вакуолей и их слиянием в центральную вакуоль, растяжением клеточных оболочек.

Рис. 5.9. Тотипотентность меристематической клетки. Производные клетки: 1 – паренхима, 2 – эпидерма, 3 – флоэма, 4 – членик сосуда ксилемы, 5 – трахеида ксилемы, 6 – склеренхимное волокно, 7 – идиобласт, 8 – колленхима, 9 – хлоренхима.
Наиболее важный момент в развитии клеток высшего растения – их дифференцировка, или специализация, то есть возникновение структурной и функциональной разнокачественности. В результате дифференцировки образуются специализированные клетки, присущие отдельным тканям. Дифференцировка осуществляется как во время растяжения, так и после окончания видимого роста клеток и определяется дифференциальной активностью генов. Дифференцировки и рост контролируется фитогормонами.
Развитие отдельных органов у растения получило название органогенеза. В целом цикле генетически обусловленное образование морфологических структур в онтогенезе называется морфогенезом. Внешние факторы, или факторы среды, также оказывают заметный эффект на рост и развитие. Свет оказывает глубокое влияние на внешнее строение растений. Свет влияет на дыхание и прорастание семян, образование корневищ и клубней, формирование цветков, на листопад, переход почек в состояние покоя. Растения, выращенные при отсутствии света (этиолированные), обгоняют в росте растения, выросшие на свету. Интенсивное освещение нередко усиливает процессы дифференцировки.
Для каждого растения существует температурный оптимум роста и развития. Температурные минимумы роста и развития в среднем лежат в интервале 5-15 ° С, оптимумы – при 35° С, максимумы – в пределах 55° С. Низкая и высокая темпера-тура может нарушать покой семян, почек, сделать возможным их прорастание и распускание. Образование цветков – это переход из вегетативного состояния в генеративное. Индуцирование (ускорение) этого процесса холодом, называют яровизацией. Без процесса яровизации многие растения (свекла, репа, сельдерей, злаки) не способны к цветению.
Огромное значение для роста, прежде всего, в фазе растяжения, имеет обеспеченность водой. Недостаток воды приводит к мелкоклеточности, отставанию в росте.
Перемещение растений в пространстве имеет ограниченный характер. Для растений свойственно, прежде всего, вегетативное движение, связанное с особенностями роста, развития и обмена веществ. Одним из примеров движения служит фототропизм – направленная реакция искривления, вызываемая односторонним освещением: при росте побеги и черешки листьев искривляются в сторону света. Многие процессы обмена веществ, роста, развития и движения подвержены ритмическим колебаниям. Иногда эти колебания следуют смене дня и ночи (циркадные ритмы), иногда связаны с длиной дня (фотопериодизм). Пример ритмических движений – ночное закрывание или открывание цветков, опускание и продольное складывание листьев, раскрытых и приподнятых в дневное время. Такие движения связаны с неравномерным тургором. Эти процессы контролируются внутренней хронометрической системой – физиологическими часами, по-видимому, существующими у всех эукариотов. У растений важнейшая функция физиологических часов – регистрация длины дня и, вместе с тем, времени года, что определяет переход к цветению или подготовку к зимнему покою (фотопериодизм). Виды, растущие на севере (севернее 60° с. ш.), должны быть преимущественно длиннодневными, поскольку их короткий вегетационный период совпадает с продолжительной длиной дня. В средних широтах (35-40° с. ш.) встречаются растения как длиннодневные, так и короткодневные. Здесь весеннее- или осеннецветущие виды относятся к короткодневным, а цветущие в разгар лета – к длиннодневным. Фотопериодизм имеет большое значение для характера распространения растений. В процессе естественного отбора у видов генетически закрепилась информация о длине дня своих местообитаний и об оптимальных сроках начала цветения. Даже у растений, размножающихся вегетативно, длина дня определяет соотношение между сезонными изменениями и накоплением запасных веществ. Виды, индифферентные к длине дня, являются потенциальными космополитами и нередко они цветут с ранней весны до поздней осени. Некоторые виды не могут выходить за пределы географической широты, определяющей их способность к цветению при соответствующей длине дня. Фотопериодизм важен и в практическом отношении, поскольку он определяет возможности продвижения южных растений на север, а северных – на юг. Одним из важных процессов, осуществляющихся в ходе индивидуального развития, является морфогенез. Морфогенез (от греческого «морфе» – вид, форма), то есть становление формы, образование морфологических структур и целостного организма в процессе индивидуального развития. Морфогенез растений обусловливается непрерывной активностью меристем, благодаря чему рост растения продолжается в течение всего онтогенеза, хотя и с разной интенсивностью. Процесс и результат морфогенеза определяются генотипом организма, взаимодействием с индивидуальными условиями развития и закономерностями развития, общими для всех живых существ (полярность, симметрия, морфогенетическая корреляция). Вследствие полярности, например, верхушечная меристема корня производит только корень, а апекс побега – стебель, листья и репродуктивные структуры (стробилы, цветки). С законами симметрии связана форма различных органов, листорасположение, актиноморфность или зигоморфность цветков. Действие корреляции, то есть взаимосвязи разных признаков в целостном организме, сказывается на характерном для каждого вида внешнем облике. Естественное нарушение корреляций в ходе морфогенеза приводит к различным тератологиям (уродствам) в строении организмов, а искусственное (путем прищипки, обрезки) – к получению растения с полезными для человека признаками.
В онтогенезе растение претерпевает возрастные изменения от эмбрионального состояния до генеративного (способного давать потомство путем образования специализированных клеток бесполого или полового размножения – спор, гамет), а затем – до глубокой старости.
Выделяют 2 группы цветковых растений по типу репродуктивных процессов: монокарпики и поликарпики. К первой группе (монокарпики) относят однолетники, часть многолетников (бамбуки), которые цветут и плодоносят только один раз в жизни. Ко второй группе (поликарпики) принадлежат многолетние травы, древесные и полудревесные растения, способные плодоносить многократно. Онтогенез цветкового растения от возникновения зародыша в семени до естественной смерти особи подразделяют на возрастные периоды – этапы онтогенеза.
1. Латентный (скрытый) – покоящиеся семена.
2. Прегенеративный, или виргинильный, – от прорастания семени до первого цветения.
3. Генеративный – от первого до последнего цветения.
4. Сенильный, или старческий, – с момента потери способности к цветению до отмирания.
В пределах этих периодов различают этапы. В группе виргинильных растений выделяют проростки (P), недавно появившиеся из семян и сохраняющие зародышевые листья – семядоли и остатки эндосперма. Ювенильные растения (Yuv), несущие еще семядольные листья, и следующие за ними ювенильные листья – более мелкие и иногда по форме еще не вполне похожие на листья взрослых особей. Имматурными (Im) считают особи, уже потерявшие ювенильные черты, но еще не вполне оформившиеся, полувзрослые. В группе генеративных растений (G) по обилию цветущих побегов, их размерам, соотношению живых и мертвых частей корней и корневищ различают молодые (G1), средневзрослые зрелые (G2) и старые генеративные особи (G3). Для высших растений очень важны процессы органогенеза. Под органогенезом понимают формирование и развитие основных органов (корня, побегов, цветков). Каждому виду растений свойствен свой темп заложения и развития органов. У голосеменных формирование репродуктивных органов, ход оплодотворения и развития зародыша достигают одного года (у ели), а иногда и больше (у сосны). У некоторых высших споровых, например у равноспоровых плаунов, этот процесс длится около 12-15 лет. У покрытосеменных процессы споро- и гаметогенеза, оплодотворения и развития зародыша происходят интенсивно, особенно у эфемеров (однолетних растений засушливых районов) – за 3-4 недели.
Для цветковых растений установлен ряд этапов органогенеза. Главнейшие из них: дифференциация стебля, закладка листьев и побегов второго порядка; дифференциация соцветия; дифференциация цветка и образование археспория в семязачатках; мега- и микроспорогенез; мега- и микрогаметогенез; зиготогенез; формирование плода и семени.
В онтогенезе организмов закономерно повторяются некоторые этапы развития, свойственные их отдаленным предкам (явление рекапитуляции). Впервые естественнонаучное объяснение рекапитуляциям дал Ч. Дарвин (1859). В 1866 г. Э. Геккель фактам повторения этапов филогенеза в онтогенезе придал форму биогенетического закона. В основе биогенетического закона лежит индивидуальное развитие особи (онтогенез), которое, в той или иной степени, представляет короткое и быстрое повторение важнейших этапов эволюции вида (филогенеза). Имеется множество примеров проявления биогенетического закона в мире растений. Так, протонема мхов, образующаяся на первых этапах прорастания споры, напоминает водоросль и свидетельствует о том, что предками мхов были, вероятнее всего, зеленые водоросли. У многих папоротников первые листья имеют дихотомическое (вильчатое) жилкование, которое было свойственно листьям ископаемых форм древних папоротников из среднего и верхнего девона. Зигоморфные цветки покрытосеменных при своем заложении проходят актиноморфную стадию. Биогенетический закон используется для выяснения особенностей филогенеза.